
⏱ 9 min read
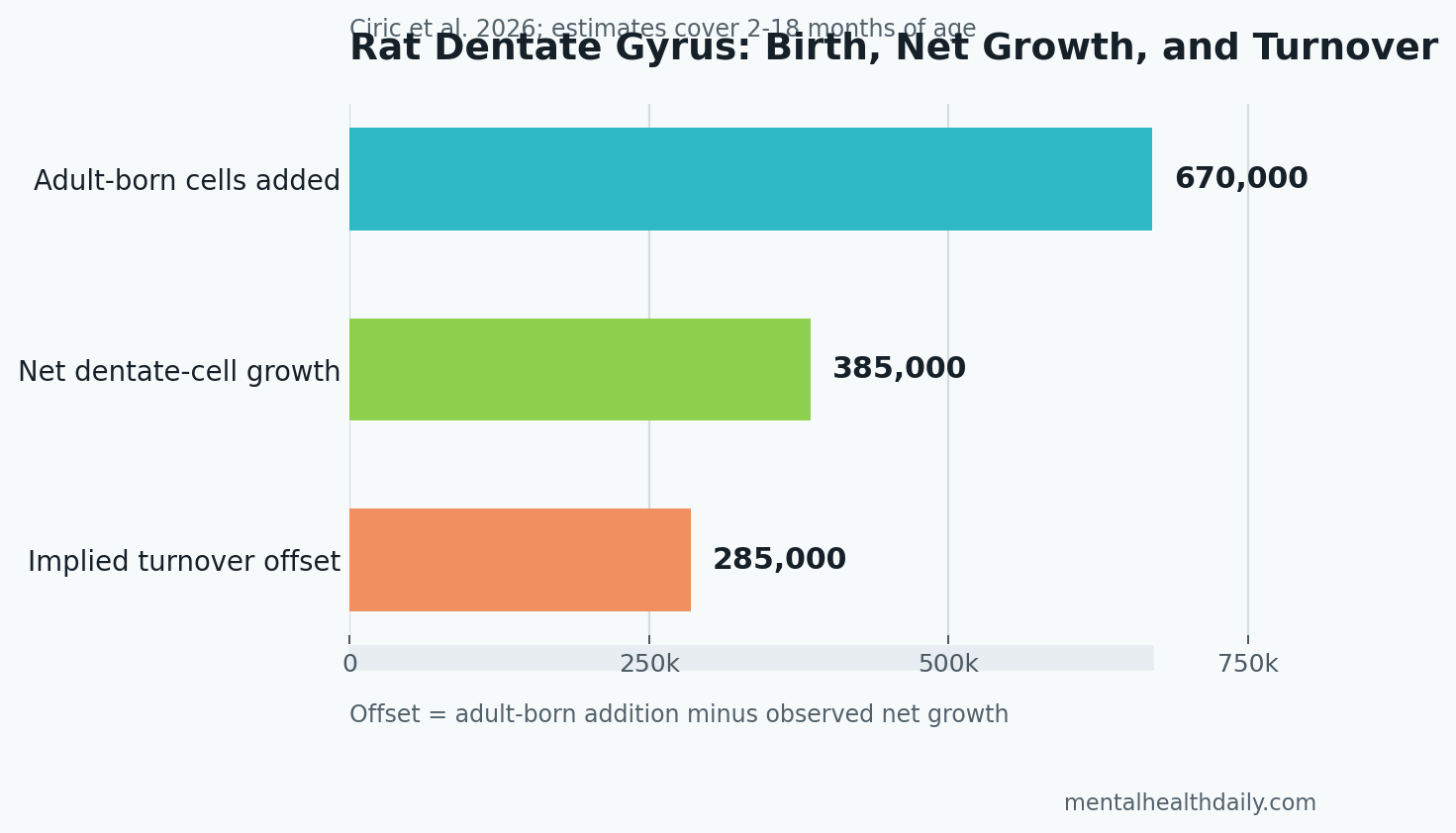
A 2026 rat study estimated that adult hippocampal neurogenesis added 670,000 dentate gyrus granule neurons from 2 to 18 months of age, but the total dentate gyrus population grew by only 385,000 cells because older developmentally born neurons were also being lost.1 The result makes adult neurogenesis look numerically larger than the usual “tiny trickle” framing, while also showing why hippocampal volume is a poor one-to-one proxy for new-neuron production.
Research Highlights
- Adult-born neurons were substantial: Ki67-based modeling estimated 670,000 adult-born dentate gyrus cells added from 2 to 18 months in male rats.1
- Net growth trailed cell birth: total dentate granule-cell number increased by 385,000 cells, a 20% rise, p = 0.003.1
- Turnover explained the gap: neurons born at postnatal day 6 declined by around 30,000 cells, or 19%, over the same adult interval.1
- Cell density increased: granule-cell layer volume decreased, R2 = 0.12, p = 0.002, while granule-neuron density increased, R2 = 0.22, p < 0.0001.1
- Human translation stays unsettled: human hippocampal neurogenesis studies disagree sharply, so this rat finding should guide mechanism, not clinical claims.23
Dentate gyrus means the hippocampal subregion that helps separate similar experiences into distinct memory traces. Adult-born neurons are new granule cells generated after early development, while developmentally born neurons are older granule cells made around birth or early life.
The specific claim is anatomical rather than a vague “the adult brain keeps growing forever” message. Ciric et al. tested a cell-accounting problem: when adult-born neurons are added and older neurons die, does the total dentate gyrus cell pool actually expand?
670,000 Adult-Born Cells Were Added but Only 385,000 Remained as Net Growth
Ciric et al. studied male Long-Evans rats sampled at 2, 4, 6, 12, and 18 months of age.1 The researchers used 3 histological readouts: Ki67 for proliferating cells, CldU for cells born on postnatal day 6, and cresyl violet stereology for total dentate granule-cell number.
Ki67 is a marker of cell proliferation. In this study, Ki67-positive cells in the subgranular zone served as the proxy for ongoing adult neurogenesis. Ki67 counts declined sharply with age, with an age effect of F4,64 = 49, p < 0.0001, and plateaued by 12 months.
The Ki67 trajectory was best fit by a cubic model, R2 = 0.75. Integrating that curve produced the headline estimate: 670,000 adult-born cells added between 2 and 18 months, assuming each Ki67-positive cell translated into one surviving adult-born granule neuron.
Total cell counting gave the calibration. Cresyl violet stereology estimated around 24,000 additional granule cells per month, or 385,000 net added cells over the same interval. The gap between 670,000 added cells and 385,000 net growth is the central finding: adult neurogenesis was large, but cell turnover masked a large share of its structural effect.
Developmentally Born Neurons Declined by 19%
CldU is a thymidine analog, a DNA-labeling chemical that marks cells dividing around the time of injection. The rats received CldU on postnatal day 6, near the developmental peak of dentate gyrus neuron production, which allowed the researchers to track long-term survival of that early-born cohort.
CldU-positive cells declined significantly with age. Linear regression estimated about 1,900 postnatal-day-6-born cells lost per month, or roughly 30,000 cells from 2 to 18 months. That was a 19% loss, R2 = 0.09, p = 0.006.1
The 30,000-cell number refers to cells born on one labeled developmental day, so it is only a sampled slice of early-born granule-cell loss. Ciric et al. proposed that if similar loss occurred across roughly 10 peak developmental days, the missing 285,000-cell difference between adult-born addition and net growth could plausibly reflect turnover of developmentally born neurons.
- Cell birth signal: Ki67 modeling suggested 670,000 adult-born cells were added.
- Total-count signal: stereology showed 385,000 net granule cells were gained.
- Turnover signal: CldU-labeled developmentally born neurons declined by 19%.
Cell Density Rose Even as the Granule-Cell Layer Got Smaller
The volume result prevents a simple “more cells equals larger structure” interpretation. Total granule-cell number increased, but granule-cell layer volume decreased from 2 to 18 months, R2 = 0.12, p = 0.002. As a result, granule-neuron density increased, R2 = 0.22, p < 0.0001.1
Stereology is a sampling method for estimating total cell number from tissue sections without simply counting every cell. That matters here because volume and density can move in different directions. A smaller-looking layer can still contain more cells if packing density increases.
Volume caution: hippocampal volume in depression, schizophrenia, electroconvulsive therapy, stress, or aging can reflect several biological layers:
- Dendritic branching or shrinkage.
- Glial, vascular, inflammatory, or extracellular-space changes.
- Cell size, cell packing, or neuron number.
New neurons are one candidate mechanism, not the whole measurement.
Older Rodent Studies Often Missed the Early Adult Window
The paper also explains why older animal studies were inconsistent. Bayer et al. reported dentate granule-cell growth from juvenile into adult rat life, with an estimate close to 1,100 cells per day.4 Ciric et al. estimated roughly 800 cells per day, but started at 2 months rather than including the juvenile interval.
Several later studies failed to find adult dentate growth. Ciric et al. argued that some of those studies began too late, used small samples, or were designed to detect age-related neuronal loss rather than early adult cell accumulation. Their own estimate put 57% of adult-born addition between 2 and 6 months, so a study beginning at 5 or 6 months could miss much of the growth window.
Rasmussen et al. studied aged rats with memory impairment and reported no loss of principal hippocampal and subicular neurons.5 Ciric et al. reanalyzed the dentate gyrus values from that work and estimated 317,000 more cells at 24 months than at 3 months, with borderline significance, p = 0.046.
The reanalysis shows how cell growth can be missed when sample size and age-window choices are not optimized for detecting it.
Human Hippocampal Neurogenesis Remains More Controversial
Human translation is the hard boundary. Spalding et al. used carbon-14 birth dating and estimated substantial adult hippocampal neurogenesis in humans.2 Boldrini et al. later reported that human hippocampal neurogenesis persisted across aging.3 Sorrells et al., by contrast, argued that human hippocampal neurogenesis dropped sharply during childhood and was undetectable in adult tissue samples.6
Those papers do not become settled by a rat dentate gyrus study. Species timing differs. Rodents produce a large share of dentate granule cells after birth, while human developmental timing may be earlier and harder to infer from adult tissue.
Best inference: the Ciric study strengthens the case that rodent adult neurogenesis can reshape dentate gyrus cell number. It does not prove that adult human hippocampi add comparable numbers of neurons or that human memory symptoms can be treated by simply increasing neurogenesis.
Depression and ECT Claims Need Extra Calibration
Hippocampal volume is often invoked in psychiatric research because depression and schizophrenia have been associated with smaller hippocampal measures, and electroconvulsive therapy can increase hippocampal volume in imaging studies.78 Neurogenesis is an appealing explanation, but the Ciric paper argues for a more complicated accounting model.
Animal work cited by Ciric et al. found that neurogenesis was not required for stress-related hippocampal volume reduction, and a 2024 electroconvulsive-stimulation study found MRI-detectable hippocampal volume increase through neurogenesis-independent mechanisms.910 Dendrites, glia, vasculature, cell size, and tissue packing can all affect volume.
Boldrini et al. did report higher hippocampal granule neuron number and dentate gyrus volume in antidepressant-treated depression cases compared with untreated cases.11 That result keeps the psychiatric question alive. The calibrated version is narrower: neuron number may contribute to some psychiatric hippocampal findings, but hippocampal volume alone cannot identify whether adult-born neurons, developmentally born neuron turnover, dendritic remodeling, or non-neuronal tissue changes drove the signal.
Memory Theories Need Turnover and Growth
The memory angle is the most interesting nonclinical implication. A dentate gyrus that only adds cells would gradually expand storage capacity. A dentate gyrus that adds adult-born neurons while removing some older neurons could support both memory formation and memory updating.
Adult hippocampal neurogenesis has been linked to forgetting in animal experiments, including work showing that increasing neurogenesis can promote loss of older hippocampus-dependent memories.12 Forgetting can be adaptive when old traces would otherwise interfere with new learning.
Turnover model: new adult-born neurons integrate into existing dentate circuits, compete for synaptic space, and may help reorganize older representations. Ciric et al. did not test behavior, so this remains a mechanism hypothesis. The cell-count data make the hypothesis anatomically plausible because addition and loss were both visible in the same system.
What This Rat Study Can and Cannot Support
Supported: in male rats, the dentate gyrus granule-cell population increased from 2 to 18 months even though developmentally born neurons declined. Adult neurogenesis was large enough to outpace measured turnover, but not large enough to make total growth equal the raw adult-born addition estimate.
Not supported: direct human memory enhancement, a depression treatment claim, or a simple claim that larger hippocampal volume equals more adult-born neurons. The study was histological and anatomical. It did not test psychiatric symptoms, memory performance, antidepressant exposure, or human tissue.
Clinical translation: hippocampal plasticity should be treated as a multi-component signal. Adult neurogenesis, developmentally born neuron survival, dendritic remodeling, inflammation, glia, and cell packing can all move the measurement. The Ciric paper makes the neuron-accounting problem clearer rather than turning it into a one-factor biomarker.
Questions About Adult Hippocampal Neurogenesis
Does this prove adults keep making new human brain cells?
No. It shows substantial dentate gyrus neurogenesis and net growth in male rats. Human adult hippocampal neurogenesis remains disputed across tissue and carbon-dating studies.
Does more adult neurogenesis always mean better memory?
No. New neurons may support flexibility and new learning, but they may also contribute to forgetting or reorganization of older traces. The useful question is which memories, which age window, and which circuit state.
Can hippocampal volume reveal neurogenesis?
No. Volume can change for many reasons besides new neurons. The Ciric study is a warning against reading imaging volume as a direct adult-neurogenesis meter.
References
- Ciric T, Cahill SP, Lin T, Choi S, Snyder JS. The dentate gyrus grows throughout life despite turnover of developmentally-born neurons. Hippocampus. 2026;36:e70099. doi:10.1002/hipo.70099
- Spalding KL, Bergmann O, Alkass K, et al. Dynamics of hippocampal neurogenesis in adult humans. Cell. 2013;153:1219-1227. doi:10.1016/j.cell.2013.05.002
- Boldrini M, Fulmore CA, Tartt AN, et al. Human hippocampal neurogenesis persists throughout aging. Cell Stem Cell. 2018;22:589-599.e5. doi:10.1016/j.stem.2018.03.015
- Bayer SA, Yackel JW, Puri PS. Neurons in the rat dentate gyrus granular layer substantially increase during juvenile and adult life. Science. 1982;216:890-892. PubMed
- Rasmussen T, Schliemann T, Sorensen JC, Zimmer J, West MJ. Memory impaired aged rats: no loss of principal hippocampal and subicular neurons. Neurobiology of Aging. 1996;17:143-147. PubMed
- Sorrells SF, Paredes MF, Cebrian-Silla A, et al. Human hippocampal neurogenesis drops sharply in children to undetectable levels in adults. Nature. 2018;555:377-381. doi:10.1038/nature25975
- McKinnon MC, Yucel K, Nazarov A, MacQueen GM. A meta-analysis examining clinical predictors of hippocampal volume in patients with major depressive disorder. Journal of Psychiatry & Neuroscience. 2009;34:41-54. PubMed
- Nordanskog P, Dahlstrand U, Larsson MR, Larsson EM, Knutsson L, Johanson A. Increase in hippocampal volume after electroconvulsive therapy in patients with depression. Journal of ECT. 2010;26:62-67. PubMed
- Schoenfeld TJ, McCausland HC, Morris HD, Padmanaban V, Cameron HA. Stress and loss of adult neurogenesis differentially reduce hippocampal volume. Biological Psychiatry. 2017;82:1-34. PubMed
- Abe Y, Yokoyama K, Kato T, Yagishita S, Tanaka KF, Takamiya A. Neurogenesis-independent mechanisms of MRI-detectable hippocampal volume increase following electroconvulsive stimulation. Neuropsychopharmacology. 2024;49:1236-1245. PubMed
- Boldrini M, Santiago AN, Hen R, et al. Hippocampal granule neuron number and dentate gyrus volume in antidepressant-treated and untreated major depression. Neuropsychopharmacology. 2013;38:1068-1077. doi:10.1038/npp.2013.5
- Akers KG, Martinez-Canabal A, Restivo L, et al. Hippocampal neurogenesis regulates forgetting during adulthood and infancy. Science. 2014;344:598-602. doi:10.1126/science.1248903
Leave a Reply