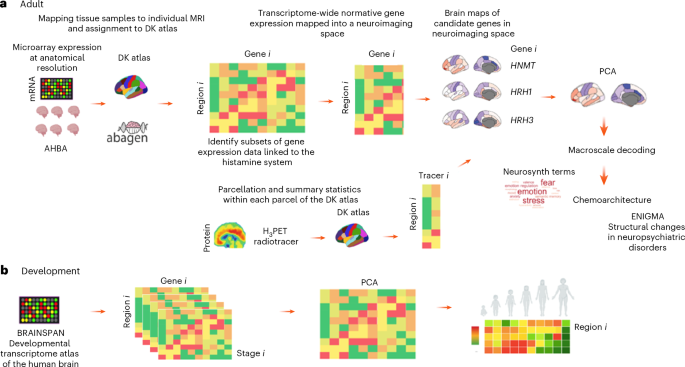
Main
Neuronal histamine plays a crucial role in the regulation of brain function, serving as a neuromodulator with widespread influence across multiple neurotransmitter systems1,2,3,4. Originating primarily from histaminergic neurons in the tuberomammillary nucleus of the hypothalamus5, histamine contributes to a wide range of physiological and cognitive processes, including arousal, attention, learning and memory, while also modulating behavioral states such as sleep–wake cycles, feeding and emotion3,6,7,8. Dysregulation of the histaminergic system has been implicated in several neuropsychiatric conditions such as anxiety, depression, schizophrenia and autism spectrum disorder, as well as neurodegenerative diseases, including Alzheimer’s, Parkinson’s and Huntington’s diseases3,9. Therefore, targeting the brain histamine system has garnered significant attention as a potential new therapeutic strategy for treating these disorders, with pharmacological interventions aimed at modulating histamine receptor activity showing promise in preclinical models9.
Histamine exerts its effects through four known histamine receptors (histamine receptor H1 (encoded by HRH1), H2 (HRH2), H3 (HRH3) and H4 (HRH4)), each of which mediates distinct functions, including neurotransmitter release, synaptic plasticity and network excitability10,11. Among them, the H3 receptor acts predominantly as an autoreceptor on presynaptic histaminergic neurons, where it tightly regulates histamine release and synthesis through a feedback inhibition mechanism12. This autoregulatory role of H3 is critical for maintaining homeostasis in histaminergic signaling as it modulates neurotransmitter release not only at histaminergic synapses but also within broader neuromodulatory networks by influencing other neurotransmitter systems, such as dopamine, serotonin and acetylcholine13. The system’s activity is regulated by two key enzymes—L-histidine decarboxylase (HDC), which synthesizes histamine from L-histidine (using a dual mechanism for translocation and degradation that ensures restriction of histamine synthesis to axonal terminals and at the same time rapid loading into synaptic vesicles14), and histamine N-methyltransferase (HNMT), which catalyzes its degradation15,16.
Comparative studies in animal models, particularly rodents17, have significantly advanced our understanding of the histamine system’s fundamental properties16,18,19,20,21,22. These studies have revealed the diverse roles that histamine plays in regulating behavior and interacting with other neuromodulatory systems23. However, notable interspecies differences in the distribution and density of histamine receptors and neurons highlight the challenges of extrapolating findings from model organisms to humans24. For example, recent studies have shown significant disparities in the spatial organization of histaminergic neurons and their projections between rodents and primates, underscoring the need for species-specific investigations16,18,19,20,21. Moreover, non-human primate studies suggest greater histaminergic involvement in higher-order cognitive functions in primates than in rodents, further emphasizing the importance of comprehensive mapping in the human brain to fully delineate the histamine system’s unique roles in complex cognition and behavior11,25.
Despite its importance, the molecular and functional architecture of the histaminergic system in the human brain remains comparatively undercharacterized relative to better-studied neuromodulatory systems such as dopamine and serotonin, for which detailed information is available on the anatomical distribution of key molecular components, interactions with other neurotransmitter systems, and system-level circuitry underlying cognition and behavior26. This relative lack of integrated, human-specific data has hindered translational progress, limiting our ability to develop targeted interventions and to understand how histaminergic dysregulation contributes to brain disorders. Notably, a substantial body of evidence has been derived from positron emission tomography (PET) and single-photon emission computed tomography studies of histamine H1 and H3 receptors in humans9,27, which has been invaluable for delineating aspects of histaminergic molecular circuitry. However, these insights have largely relied on smaller or heterogeneous datasets and have only occasionally been integrated with functional measures, pharmacological challenges or broader neurobiological context to assess target engagement or system-level relationships. As a result, a comprehensive, multiscale mapping of the histamine system in the human brain remains a critical unmet need. Recent advances in transcriptomic profiling and neuroimaging now enable the investigation of the spatial organization of histamine-related genes in the human brain at unprecedented resolution. Crucially, these approaches allow the integration of molecular data—such as gene expression and receptor distribution—with functional and structural brain imaging, offering new opportunities to elucidate how histaminergic signaling shapes neural circuits, cognition and behavior.
By employing an integrative multimodal approach, this study provides a comprehensive analysis of the genetic, molecular and functional architecture of the histamine system in the human brain (see Fig. 1 for a summary of our pipeline). Using regional transcriptomic data and single-cell RNA sequencing data from the Allen Human Brain Atlas (AHBA), alongside in vivo molecular imaging normative templates28, we examine the spatial distribution of histamine-related genes and their spatial covariance with the distribution of molecular targets of neurotransmitter systems. These data can provide a descriptive anchor to better understand existing collected data (for example, cellular and regional distribution mapping, time courses) but also generate hypotheses alongside, for example, regarding brain processes for which the histamine system might contribute based on analyses of regional patterns of co-distribution (for example, high expression of elements of the histamine machinery in regions highly important for a specific cognitive process or brain disorder). Therefore, in addition, we examined how the spatial distribution of histaminergic gene expression and meta-analytical maps of cognitive functions/behavior correlate with structural brain changes in the main psychiatric disorders to generate hypotheses about potential functional implications and relevance to mental health. Finally, we characterized the developmental trajectories of histaminergic brain gene expression7, shedding light on how this system matures across critical periods of brain development.

Summary of the analytical pipeline and data sources. a, Adult transcriptomic data. b, Developmental transcriptomic data. DK, Desikan–Killiany; MRI, magnetic resonance imaging.
Results
Cellular distribution of histaminergic gene expression
Single-nucleus RNA sequencing data revealed distinct expression patterns of histaminergic genes across major cortical cell types. HNMT, which encodes the key enzyme for histamine degradation, exhibited a widespread distribution, with significant expression in both neurons and non-neuronal cells. Aldehyde dehydrogenase enzyme 7 family member A1 (ALDH7A1) was highly expressed by astrocytes and monoamine oxidase B (MAOB) in excitatory neurons. Among histamine receptors, HRH3 demonstrated the most neuronally restricted profile, predominantly expressed in inhibitory neurons, while HRH1 and HRH2 displayed broader neuronal expression, predominating in excitatory neurons. Data were not available for HRH4, HDC and AOC1. These findings establish a cellular framework for histamine signaling in the human cortex (Fig. 2).

The heatmap displays normalized single-cell mRNA expression of histamine-related genes, normalized to highlight variation across main cell clusters. Gene expression was measured in multiple cortical areas of the human brain using RNA sequencing. Data are available as part of the Allen Human Brain Cell Atlas18 and can be downloaded from ref. 105. OPC, oligodendrocyte precursor cells; VLMC, vascular leptomeningeal cells.
Regional expression of histaminergic genes in the adult human brain
Postmortem microarray data from the AHBA revealed heterogeneous regional expression of histaminergic genes across the brain. HRH1 showed relatively high expression in the frontal cortex and limbic structures such as the entorhinal, amygdala, parahippocampal cortices and cingulate gyrus. HRH2 is more variably expressed, with lower levels in the limbic system and cingulate cortex but relatively higher expression in some primary sensory regions such as the cuneus and lateral occipital cortex. HRH3 has lower-than-average expression in occipital and parietal cortices but is highly expressed in frontal regions, anterior cingulate and subcortical regions such as the caudate, putamen and thalamus. HRH4 has more localized expression, with moderate levels in the middle cingulate, entorhinal cortex, hippocampus and amygdala. The enzymes involved in histamine metabolism also show region-specific expression. HDC, which synthesizes histamine, is particularly expressed in limbic regions such as the entorhinal and parahippocampal cortices, amygdala and hippocampus. HNMT, responsible for histamine degradation, is broadly expressed but with lower-than-average levels in visual and occipital regions and higher-than-average expression in the orbitofrontal and middle cingulate, hippocampus and amygdala. MAOB, which encodes another enzyme contributing to histamine metabolism, is highly expressed in subcortical structures such as the putamen and pallidum, orbitofrontal cortex and cingulate. ALDH7A1, linked to histamine catabolism, exhibits strong expression in the middle cingulate, thalamus and basal ganglia (Fig. 3).

Surface-rendered cortical and subcortical maps show regionally normalized (Z-scored) expression of key histamine-related genes (HDC, HNMT, ALDH7A1, MAOB, HRH1–HRH4) across the human brain. For each gene, lateral and medial cortical views and subcortical projections are displayed. PCA of regional gene expression profiles captures the dominant axis of histaminergic molecular variation (top right). Bottom panels show normative PET templates of histamine H3 receptor binding derived from two independent radiotracers, [11C]GSK-189254 and [11C]MK-8278, displayed as Z-scored regional distributions. The heatmap summarizes Spearman correlations between regional PET binding patterns and gene expression maps (including PCA scores). Asterisks indicate statistically significant correlations after correction for spatial autocorrelation (spin rotation permutation two-sided tests). Color bars indicate Z-score ranges (blue represents lower relative expression or binding; red represents higher relative expression or binding).
Principal component analysis (PCA) on histaminergic gene expression identified a dominant spatial component that captured shared variance across histamine-related genes, reflecting a functional gradient from primary sensorimotor areas to multimodal association regions. This first component alone explained 41.1% of the total variance (Supplementary Fig. 1). The loadings of each gene into PCA1 can be seen in Fig. 4. Except for HRH2, which loaded negatively, all other genes had positive loadings. PCA1 scores were higher and lower than chance, respectively, in the limbic and visual 7 Yeo networks. For the von Economo cytoarchitectonic classes, we found PCA1 scores to be higher and lower than chance, respectively, in the limbic and primary sensory cortices (Supplementary Fig. 2).

The left panel shows Spearman correlations between the regional histaminergic molecular score (derived from the PCA of histamine-related gene expression) and normative PET templates of major neurotransmitter receptors and transporters across the human brain. Colors indicate correlation strength (red = positive, blue = negative), and asterisks denote associations surviving false discovery rate (FDR) correction for multiple comparisons. Right panels summarize cognitive and behavioral domains significantly associated with the histaminergic molecular axis using Neurosynth meta-analytic decoding. Word clouds depict terms showing significant positive (top) and negative (bottom) correlations after FDR correction, with word size reflecting the relative strength of association. Positive associations preferentially map onto affective, stress-related and emotion regulation processes, whereas negative associations are enriched for visual, attentional and perceptual functions. Significance was assessed using spin rotation two-sided permutation tests.
Spatial correlations with H3 receptor binding and neurotransmitter systems
To validate our inferred transcriptomic signature, we compared histaminergic gene expression maps with in vivo PET imaging of H3 receptor distribution. Regional H3 receptor bindings showed the highest positive significant correlation with HRH3 expression but also correlated positively with HRH1, MAOB and PCA1 scores, and negatively with HRH2 (Table 1 and Fig. 3). For [11C]MK-8278 only, we also found significant positive correlations with the expression of HNMT. These findings support our assumption that mRNA expression data might be a good proxy mapping of the architecture of the human brain histamine system.
In addition, PCA-derived histaminergic expression patterns correlated with receptor densities of multiple neurotransmitter systems. Notably, the spatial distribution of histaminergic PCA1 scores positively correlated with the one for serotonin receptors 5-HT1A and 5-HT2A, dopamine receptor D1 and mu opioid receptor, although only the first survived correction for multiple comparisons. Moreover, it negatively correlated with the distribution of serotonin receptors 5-HT1B, 5-HT4 and 5-HT6, dopamine transporter and dopamine D2 receptor, nicotinic receptor A4B2, muscarinic receptor M1 and acetylcholine transporter, noradrenaline transporter, cannabinoid receptor 1, and glutamate N-methyl-d-aspartate (NMDA) and mGlu5R receptors (Table 2 and Fig. 4). We did not find any significant correlation between the spatial distribution of PCA1 scores and any of the six magnetoencephalography (MEG) cortical maps of oscillatory power across different bands (Supplementary Table 1 and Supplementary Fig. 3).
Macroscale functional decoding of histaminergic gene expression
To explore the functional implications of histaminergic gene expression, we correlated PCA-derived histaminergic expression patterns with meta-analytic functional neuroimaging maps from Neurosynth. Brain regions with higher histaminergic gene expression were significantly associated with brain response during tasks indexing the following cognitive domains related to emotion, emotion regulation, stress, fear and anxiety processing, salience detection, impulsivity and cognitive control, loss, risk and addiction, sleep and memory, eating and pain. Brain regions with lower-than-average histaminergic gene expression were significantly associated with visual function, attention, gaze, multisensory integration, mental imagery and reading (Fig. 4 and Supplementary Table 1).
Histaminergic gene expression and neuropsychiatric disorders
Histaminergic gene expression patterns were next examined in relation to structural brain alterations observed in major neuropsychiatric disorders. Using ENIGMA consortium data, we identified significant correlations between PCA-derived histaminergic expression and disorder-specific cortical alterations in thickness and surface area. PCA1 scores showed the strongest negative associations with thickness changes in attention deficit hyperactivity disorder (ADHD) and major depressive disorder (MDD), and a significant positive association with thickness changes in anorexia nervosa. For the surface area, we found significant positive associations with the spatial distribution of changes in anorexia nervosa and schizophrenia (Supplementary Fig. 4).
Developmental trajectories of histaminergic gene expression
Analysis of the BrainSpan transcriptomic developmental atlas revealed dynamic changes in histaminergic gene expression across lifespan stages, with distinct regional time courses likely shaping histamine-related functions. HDC expression peaked during late fetal development and early postnatal stages, consistent with the early establishment of histaminergic neurotransmission. HRH3 exhibited a protracted developmental trajectory, increasing steadily from childhood to adulthood, mirroring the maturation of prefrontal cognitive control networks. HNMT expression remained relatively stable across development, probably reflecting its housekeeping role in histamine metabolism (Fig. 5).

The heatmap displays whole-brain normalized gene expression across developmental stages. Data were extracted from the BrainSpan Atlas of the Developing Human Brain (https://www.brainspan.org/). Significance was assessed using permutation testing where we shuffled developmental stages across samples. The * denotes P < 0.05 (two-sided) after FDR correction for the number of genes. RPKM, reads per kilobase of transcript per million mapped reads.
We also mapped regional time courses of histaminergic gene expression and identified region-specific maturation patterns of PCA1 scores summarizing developmental histaminergic brain gene expression. PCA1 explained 66.10% of the variance and had negative loadings from all candidate genes. We found PCA1 scores to follow a delayed trajectory in association cortices, reaching peak expression in late adolescence, whereas its expression in primary sensorimotor regions stabilized earlier. Moreover, it showed early enrichment in subcortical structures, reinforcing histamine’s role in early neuromodulation (Supplementary Fig. 5).
Discussion
This study provides an integrated characterization of the histaminergic system in the human brain, leveraging transcriptomic, neuroimaging and functional datasets to delineate its molecular organization and relevance to brain function underlying cognition and psychiatric disorders. The findings reveal that histaminergic genes exhibit distinct cellular and regional expression profiles, closely aligning with known histaminergic neuroanatomy and function16,18,19,20,21. HRH1 and HRH2 show the highest expression in excitatory neurons, reinforcing histamine’s role in modulating cortical excitability4,29. By contrast, HNMT, which degrades histamine, is more broadly expressed, highlighting its role in shaping histaminergic tone across multiple brain regions. The receptor profiles further underscore functional specialization within the histaminergic system, with HRH3 exhibiting a neuronally restricted distribution in inhibitory circuits, suggesting a central role in feedback regulation3,23. Although single-nucleus RNA sequencing revealed enrichment of HRH3 expression within inhibitory neuronal populations, this finding should be interpreted in light of the methodological scope of transcriptomic data, which cannot resolve presynaptic versus postsynaptic localization or distinguish local interneurons from long-range projecting cells. The H3 receptor serves dual roles as both an autoreceptor and heteroreceptor, modulating histamine release as well as the release of other neuromodulators, and these functions are not incompatible with transcriptional enrichment in inhibitory neurons12. Rather, this pattern may reflect a role for H3 receptors in regulating cortical excitability via inhibitory microcircuits. Consistent with this interpretation, regional H3 receptor binding measured using PET showed strong correspondence with HRH3 gene expression, supporting the relevance of the transcriptomic findings despite cell-type-level ambiguities30.
At the regional level, our summary histaminergic gene expression signature follows a heterogeneous spatial distribution, supporting the notion of a functional specialization across the brain. HRH1 is prominently expressed in the frontal cortex and limbic regions, such as the entorhinal cortex, amygdala and cingulate gyrus, highlighting its putative role in emotional regulation and higher-order cognitive functions31,32. By contrast, HRH2 shows relatively higher expression in primary sensory areas such as the cuneus and lateral occipital cortex, suggesting a putative role in sensory processing. HRH3 exhibits strong expression in subcortical regions, including the caudate, putamen and thalamus, as well as frontal areas, consistent with its role in modulating motor control, reward processing and neurotransmitter release33. HRH4 is more localized, with moderate presence in limbic regions such as the hippocampus, amygdala and entorhinal cortex, aligning with its putative role in the brain’s immune responses as these regions are also known to contain a higher density of cells with immune function34. Enzymes regulating histamine metabolism also show distinct regional expression: HDC, responsible for histamine synthesis, is particularly active in limbic areas; HNMT, involved in histamine degradation, is broadly expressed in the brain with higher levels in the orbitofrontal cortex, hippocampus and cingulate; and MAOB and ALDH7A1, which are also involved in histamine metabolism, exhibit strong expression in subcortical and cingulate regions. Together, these patterns hint at the possibility of diverse and region-specific roles of histaminergic signaling in sensory processing, cognition, emotion and motor control, with implications for understanding its involvement in both normal brain function and neuropsychiatric disorders35,36.
The validation of the inferred transcriptomic signature through comparison with in vivo PET imaging of H3 receptor distribution as measured using two different tracers in two different cohorts provides critical support for our spatial transcriptomic mapping of the human brain’s histaminergic system. Although also expressed postsynaptically, including in the striatum, being an autoreceptor expressed in histaminergic presynaptic terminals, H3 receptor binding is likely to offer a reliable proxy for histaminergic innervation13. The strong positive correlations between regional H3 receptor binding and HRH3 expression highlight the congruence between mRNA expression and receptor localization, confirming the robustness of transcriptomic data as a reliable spatial representation of histaminergic architecture.
Whether the transcriptomic signature identified here also predicts regional variation in histamine H1 receptor availability remains an open empirical question. While human H1 receptor PET studies exist, they are currently limited to a small number of cohorts and are not yet available as harmonized, shareable whole-brain templates suitable for large-scale multimodal integration. Future efforts aimed at generating standardized H1 receptor PET templates—ideally with prospective attention to data sharing and cross-site harmonization—would therefore represent a valuable extension of the present work. Notwithstanding these limitations, existing clinical and imaging evidence underscores the translational relevance of histaminergic signaling across neuropsychiatric conditions, including depression, anorexia nervosa, schizophrenia and aging-related neurodegeneration. In particular, altered H1 receptor binding has been reported in individuals with anorexia nervosa, implicating histaminergic mechanisms in the regulation of appetite, arousal and affective processing37,38. In this context, the positive association we observed between the principal component summarizing histaminergic gene expression and brain alterations in anorexia nervosa provides convergent, systems-level support for a role of histamine in eating disorders. Although the clinical literature remains relatively sparse, recent work highlights histamine’s involvement in visceral processing, stress responsivity and mood regulation in this population, encouraging further preclinical investigation and the exploration of histaminergic targets in future clinical studies39.
The positive correlation of histaminergic PCA1 scores with serotonin receptor 5-HT1A speaks in favor of histamine’s role in regulating serotonergic transmission and aligns well with preclinical evidence showing that histaminergic neurons project to the raphe nuclei, where H3 receptor activation reduces serotonin release, potentially influencing mood and cognition23,40,41. We note however that even if, for example, the H3 receptor has been found to co-exist in heteromers with the dopamine receptor D142, NMDA receptors42 and the adenosine receptor A143, to date there has been no reports of heteromers with the 5-HT1AR, and our data did not specifically show significant positive correlations with D1 or NMDA binding (which was strongly anticorrelated). Nevertheless, we should highlight that even if not reaching significance in our strict spin test, D1 was among the targets with the strongest positive correlations, an observation that urges some caution in excluding the presence of heteromers on the basis of our data exclusively. The observed negative correlations in receptor expression between the histaminergic system and other neuromodulatory systems (for example, serotonin other than 5-HT1A, dopamine other than D1 receptors, glutamate, acetylcholine) is interesting as it suggests there are region-specific specializations and/or feedback regulation in the gene expression of these neuromodulatory systems11,23. Supporting the later interpretation, histamine can suppress dopamine and glutamate release via H3 receptor-mediated inhibition while simultaneously enhancing dopamine output through H1 receptor activation40,44,45. It also modulates norepinephrine release and facilitates cholinergic transmission through projections to key regions such as the prefrontal cortex, striatum and basal forebrain40,46,47. Thus, our findings extend this preclinical evidence to suggest similar interactions may occur in humans. We interpret the inverse patterns of receptor expression as representing regulatory feedback, whereby elevated histaminergic tone reduces the necessity for receptor upregulation in other systems to preserve neurotransmitter balance. While this interpretation needs testing in humans, collectively, these findings underscore the potential integrative role of histamine as a dynamic regulator of multiple neuromodulatory pathways, wherein functional interactions are shaped by system-level feedback.
The correlation between PCA-derived histaminergic gene expression patterns and meta-analytic functional neuroimaging maps from Neurosynth indicates that brain regions with higher histaminergic gene expression were strongly associated with cognitive domains and processes related to emotion, emotional regulation, stress, fear and anxiety processing, salience detection, impulsivity, cognitive control, risk-taking, addiction, sleep, memory, eating and pain. These findings provide evidence for a role of the histaminergic system in linking physiological arousal (which leads to histamine release in the brain) with higher-order cognitive and affective processes48. The associations with stress, anxiety and emotional regulation extend previous rodent evidence for histamine’s role in adapting to environmental challenges and maintaining emotional homeostasis49 to indicate this is the case in humans. The link to salience detection and decision-making suggests histaminergic involvement in facilitating the prioritization of relevant stimuli and modulating cognitive control50. Furthermore, the strong associations with impulsivity, addiction and risk-taking behaviors highlight histamine’s contribution to reward processing and behavioral regulation within circuits governing motivation and decision-making under uncertainty49,51. The connections to sleep52, eating53 and pain54 further suggest a broader role for histamine in managing homeostatic processes, supporting its involvement in maintaining the body’s internal balance and response to environmental demands. Together, these results provide an integrated view of how histaminergic signaling supports the brain’s ability to navigate affective, cognitive and physiological challenges, making it a good candidate for a critical regulator of brain physiology in both health and neuropsychiatric disorders11,55.
The potential relevance of histaminergic signaling to neuropsychiatric disorders is further underscored by its associations with structural brain changes observed in conditions such as schizophrenia, depression, ADHD and anorexia. PCA1 scores, representing an aggregate histaminergic gene expression pattern, correlated positively with cortical thickness alterations in anorexia nervosa but negatively with thickness changes in ADHD and MDD. The negative correlation between PCA1 and cortical thinning in ADHD aligns well with the idea that disrupted histamine signaling may contribute to attentional and cognitive dysfunction56. This is consistent with preclinical studies showing that histamine depletion leads to impairments in attention and executive function57. The evidence linking histamine to depression is manifold. Chronic treatment with pitolisant, an inverse H3 agonist, was shown to prevent the development of anxiety-like behaviors in a rodent model of corticosterone-induced depression58. Selective serotonin reuptake inhibitors, which constitute first-line treatments for the treatment of depression, were shown to require an intact histaminergic system to produce therapeutic effects in a depression-like rodent model59. In addition, increased brain histamine has been shown to likely mediate suppression of SSRI-induced increases in serotonin during acute inflammation60, offering a new angle to our understanding of treatment-resistant depression often observed in patients with low-grade chronic inflammation61. In terms of surface area changes, PCA1 scores showed a positive correlation with alterations in anorexia and schizophrenia. These findings align with previous evidence implicating the histaminergic system in schizophrenia’s pathophysiology, particularly its role in modulating neurotransmitter systems such as dopamine and glutamate, both of which are dysregulated in the disorder62,63,64. Preclinical and clinical trial data have also demonstrated the therapeutic potential of targeting histamine H2 and H3 receptors in improving both cognitive dysfunction and negative symptoms in schizophrenia29,65. For example, H3 receptor antagonists, such as pitolisant, have shown promise in enhancing cognitive performance in animal models of psychosis, with early clinical trials in humans showing tolerability and potential improvements in attention and memory35,66. In addition, the positive association between PCA1 scores and cortical thickness changes in anorexia nervosa could reflect the interaction of histamine with brain areas implicated in feeding behavior, anxiety and reward processing. Evidence from studies in patients with anorexia suggests altered histamine signaling may play a role in dysregulated appetite, stress response and emotional processing8. Future clinical trials targeting histaminergic pathways may provide valuable insights into treatments for psychiatric conditions characterized by altered neurotransmitter dynamics and associated structural brain changes55,67.
Histaminergic gene expression follows a dynamic developmental trajectory, with HDC expression peaking during fetal development and early postnatal stages, reinforcing histamine’s role in early neural circuit formation. Interestingly, the protracted increase in HRH3 expression from childhood to adulthood mirrors the maturation of prefrontal cognitive control networks, which aligns well with the view that histamine plays a role in shaping executive function. Developmental analyses using regional data further demonstrate that histaminergic gene expression follows distinct regional time courses, with early maturation in sensorimotor regions and delayed expression in association cortices. This extended developmental trajectory aligns with the putative histamine’s involvement in cognitive flexibility and learning, functions that continue to mature into adulthood2,7. Together, these findings suggest that histaminergic signaling might play a fundamental role in neurodevelopmental processes and cognitive maturation, making it a candidate for further longitudinal developmental studies in clinical populations examining, for example, whether functional genetic variation in genes of the histamine pathway might shape trajectories of brain development from early infancy into adulthood.
Although this work places particular emphasis on the histamine H3 receptor—motivated by its role as a presynaptic auto- and heteroreceptor and by the availability of harmonized in vivo PET templates—the histaminergic system cannot be fully understood without considering the broader contributions of H1 and H2 receptors. Indeed, the modern study of brain histamine emerged from early anatomical and pharmacological efforts to identify histaminergic neurons and characterize H1 receptor binding in the vertebrate brain, followed by the development of PET radiotracers such as [11C]doxepin68 that enabled the first in vivo mapping of histamine receptors in humans. This foundational body of work established histamine as a key modulator of arousal, attention and cortical excitability, providing an essential framework for subsequent molecular and systems-level investigations. Consistent with this historical literature, our transcriptomic analyses revealed that HRH1 and HRH2 expression is preferentially enriched in excitatory neuronal populations, particularly within frontal and limbic cortices. This pattern aligns with experimental evidence demonstrating that H1 and H2 receptor activation enhances neuronal excitability, modulates glutamatergic transmission and influences large-scale cortical state regulation. Although we were unable to directly validate these findings using in vivo H1 or H2 receptor PET data—due to the absence of publicly available, harmonized whole-brain templates—the observed cellular and regional expression profiles provide an important molecular substrate for interpreting previous clinical and pharmacological observations. Notably, alterations in H1 receptor availability and function have been reported across a range of neuropsychiatric and neurological conditions37, including depression69, anorexia nervosa, schizophrenia62, aging and Alzheimer’s disease70,71. PET studies using H1 receptor ligands and pharmacological challenge paradigms have linked histaminergic dysregulation to changes in arousal, mood, cognitive performance and sedation-related side effects of psychotropic medications72. In anorexia nervosa in particular, reduced H1 receptor binding has been associated with altered cortical and limbic processing, offering a potential mechanistic bridge between histamine signaling, interoceptive processing and affective regulation. Within this context, the association we observed between a principal component summarizing histaminergic gene expression and disorder-specific cortical alterations provides convergent, systems-level support for the relevance of histamine pathways in eating disorders, despite the current lack of direct multimodal H1 receptor validation. By contrast, the H3 receptor occupies a distinct regulatory position within the histaminergic system, acting primarily as a presynaptic modulator of histamine synthesis and release while also regulating the release of other neurotransmitters, including dopamine, norepinephrine and acetylcholine. The strong spatial correspondence we observed between HRH3 gene expression and H3 receptor PET binding, across independent tracers and cohorts, supports the use of H3 receptor imaging as a systems-level proxy for histaminergic innervation. Importantly, this validation strengthens confidence in the broader transcriptomic signature identified here, including components driven by HRH1 and HRH2, even in the absence of direct receptor-specific imaging for these targets. Finally, it is important to note that histamine H2 receptor imaging remains largely unexplored in humans as no validated PET radiotracers are currently available for reliable whole-brain in vivo assessment. This represents a significant methodological gap in the field and limits the ability to directly test hypotheses regarding H2-mediated mechanisms in cognition and psychiatric disorders. The development of selective, shareable H1 and H2 receptor PET ligands—together with prospective attention to data harmonization and open sharing—would therefore constitute a major advance, enabling future studies to extend integrative multimodal frameworks such as the one presented here and to more fully disentangle receptor-specific contributions to histaminergic function in health and disease.
While this study offers a comprehensive characterization of the histaminergic system, several limitations should be considered. First, and foremost, our findings are correlational in nature and should be interpreted as hypothesis-generating for further experimental work rather than direct proofs. To achieve this goal, future work relying on drug challenges and longitudinal imaging genetics designs in both healthy controls and patients with neuropsychiatric disorders will be pivotal to clarify how histaminergic signaling trajectories shape brain maturation and psychiatric risk. One potential example would be pharmacological studies with H3 receptor antagonists such as pitolisant, which are known as inducers of synaptic histamine release and would allow testing the effects of targeted release of histamine in the brain on behavior and cognition in both healthy and clinical populations73. Second, the reliance on postmortem transcriptomic datasets, although valuable for spatial mapping, does not capture dynamic and diurnal fluctuations in histamine signaling that occur in vivo74,75. The integration of PET imaging with gene expression data strengthens our findings, but PET measures of receptor binding provide indirect estimates of functional histaminergic activity rather than real-time neurotransmission75. Future studies incorporating functional imaging techniques with drug challenges could offer a complementary assessment of histamine’s role in living human brains. Third, the relatively small sample sizes of several source datasets—especially those informing histaminergic gene expression—limit statistical power and may reduce the generalizability of the observed spatial and molecular associations. For example, the AHBA derives from six donors only, and data from the right hemisphere was collected for only four donors, which raises issues regarding the capacity to appropriately capture interindividual variability76. The same applies regarding sex (only one donor is female) and age (24–57 years) representation. Although the AHBA remains as the sole most anatomically detailed of the transcriptomics of the human brain, it is clear there is much to be done on diversity and inclusion to ensure representativeness measures to current standards of human neuroscience. Fourth, it is important to note the resolution constraints of microarray and RNA sequencing data, which do not fully capture cellular heterogeneity within complex brain regions. Fine-grained single-cell and spatial transcriptomic approaches could refine our understanding of histaminergic gene expression at a more granular level using spatial single transcriptomics to dissect potential differences among, for example, cortical layers and regional microcircuitry24,77. Given histamine’s widespread neuromodulatory effects, a deeper exploration of histamine’s interactions with other neurotransmitter systems may yield insights into cross-system regulation and homeostatic balance. This could be achieved by leveraging recent advances in multiscale large biophysical modeling approaches integrating transcriptomics, receptor binding, neurotransmitter dynamics and computational simulations to help bridge the gap between molecular findings and systems-level histaminergic function, ultimately providing a more mechanistic understanding of its role in cognition and neuropsychiatric disorders78,79.
Conclusion
To conclude, this study offers a foundational systems-level multimodal mapping of the histaminergic system in the human brain, integrating molecular, functional and developmental perspectives. By linking histaminergic gene expression to brain-cell types, neurotransmitter systems, cognitive domains and psychiatric disorders, these correlational findings generate several hypotheses concerning histamine’s critical role in brain organization, neurodevelopment and mental health, which further experimental mechanistic work should prioritize and build onto investigate causal relationships.
Methods
Overview of analytical approach
Briefly, our pipeline followed a structured approach to analyzing the expression of genes related to the histamine system in the human brain, focusing on both adult and developmental stages. The analysis focused on key genes involved in neuronal histamine metabolism, signaling and interactions with other neurotransmitter systems. We examined HDC, the sole enzyme responsible for histamine synthesis, and HNMT, the primary enzyme for histamine degradation in the brain. In addition, we included AOC1, MAOB and ALDH7A1, which contribute to alternative histamine degradation pathways. We also analyzed the four known histamine receptors (H1, H2, H3 and H4 receptors), which mediate histamine’s diverse neuromodulatory effects. H1 and H2 receptors regulate excitatory neurotransmission, cognition and synaptic plasticity, while H3 receptor acts as a presynaptic autoreceptor, modulating histamine release and influencing dopamine, serotonin and acetylcholine systems. H4 receptor, although primarily linked to immune function, has emerging relevance in neuroinflammation and brain disorders80,81,82.
In the adult analysis, we used microarray gene expression data from the AHBA83 to map regional variation in gene expression in neuroimaging space. We used the abagen toolbox84 to assign these data to specific brain regions on the basis of the DK atlas85, which was selected for several practical and conceptual reasons. Critically, the AHBA does not provide a native, donor-consistent anatomical parcellation in neuroimaging space. Instead, it consists of sparsely and irregularly distributed tissue samples whose spatial locations and coverage vary substantially across donors. As such, mapping AHBA microarray expression data to a standardized brain atlas is necessary to enable regional aggregation, cross-donor normalization and spatial alignment with in vivo neuroimaging data. The DK atlas offers a widely used, anatomically grounded parcellation with 83 cortical and subcortical regions defined in standard neuroimaging space. Its native implementation within the abagen toolbox facilitates reproducible preprocessing choices, including probe selection, donor-wise normalization and region-level aggregation, while also allowing robust handling of inter-hemispheric asymmetries and uneven sampling density across donors. Importantly, the use of the DK atlas ensures spatial compatibility across all multimodal datasets integrated in this study. PET receptor density templates derived from the neuromaps framework, meta-analytic functional maps from Neurosynth, and MEG-derived oscillatory power maps were either natively available or most reliably parcellated in DK space. This common anatomical framework enables direct spatial correspondence across transcriptomic, molecular imaging and functional datasets without requiring additional interpolation or cross-atlas registration steps, thereby improving reproducibility and interpretability. While higher-resolution parcellations (for example, Glasser and Schaefer atlases) offer finer spatial granularity, they are optimized primarily for functional connectivity and can be more susceptible to noise, partial volume effects and sampling bias when applied to sparse postmortem transcriptomic data. Given our goal of integrating gene expression with molecular imaging and large-scale functional decoding, the DK atlas provided the most balanced and tractable solution.
Next we applied PCA to extract dominant patterns in their expression. Since we relied on gene expression to infer regional variation in the molecular architecture of the histamine system in the human brain, which does not necessarily reflect protein levels, we used PET data from the regional variation in H3 receptor binding in normative samples of healthy participants86 to examine spatial correlations with the expression of candidate genes and the scores of the first principal component summarizing gene expression. While a bulk of data exists from efforts mapping human brain H1 receptors mostly using [11C]doxepin27,87, we could not access any suitable template data to be incorporated in our analyses. Therefore, while we discuss findings using this tracer, its omission from our current pipeline mostly reflects restraints on data sharing that made it impossible to test how well our transcriptomics signature would align against a normative signature of the H1 receptor protein as assessed by PET. This similarly explains its absence from Supplementary Table 1, where we include only tracers for which we had accessible template data. Since histamine also modulates the release of other neurotransmitters, we used PET templates of the binding of multiple receptors in the living human brain to assess the spatial correlation between regional scores of the first PCA of gene expression and parcellated data of the chemoarchitecture of the human brain28. To establish functional relevance, we examined the spatial correlation between the first principal component and meta-analytical maps of terms pertaining to different cognitive functions and behavioral domains using data from the Neurosynth repository88. Finally, we also investigated the potential relevance to neuropsychiatric disorders by assessing spatial correlations between regional PCA1 scores and structural brain changes using ENIGMA consortium data89. In the developmental analysis, we used data from the BrainSpan Atlas90 to examine histaminergic gene expression across different stages of brain development. We applied PCA to summarize developmental trajectories, capturing global and regional dominant trends in histaminergic gene expression over time (Fig. 1).
Data sources
Single-nucleus RNA sequencing data
We used single-nucleus transcriptomes from 49,495 nuclei across multiple human cortical areas available as part of the Allen Human Brain Cell Atlas. The methods underlying the generation of this dataset have been described in detail elsewhere91. Briefly, individual layers of cortex were dissected from tissues covering the middle temporal gyrus, anterior cingulate cortex (also known as the ventral division of medial prefrontal cortex, A24), primary visual cortex, primary motor cortex, primary somatosensory cortex and primary auditory cortex derived from the human brain. Nuclei were dissociated and sorted using the neuronal marker NeuN. These regions represent the full extent of currently available human single-nucleus transcriptomic data with layer-specific dissection and standardized NeuN-based neuronal sorting in this resource. They were selected by the original dataset authors to maximize anatomical and functional diversity across sensory, motor, association and paralimbic cortices rather than by the present study. Nuclei were sampled from postmortem and neurosurgical (middle temporal gyrus only) donor brains, and expression was profiled with SMART-Seq v4 RNA sequencing. Normalized gene expression was downloaded from ref. 92. To summarize gene expression in the main brain-cell types, we averaged expression of candidate genes across cells of the same cluster91.
Regional microarray expression data
Gene expression data from the AHBA were obtained from six adult postmortem brains aged 24–57 years. Genetic probes were reannotated following standardized methods outlined in ref. 93 to increase accuracy, and probes deemed unreliable were excluded. For each gene, the most stable probe was selected on the basis of pooled donor correlations, resulting in a final dataset of 15,633 probes across the brain. Microarray samples were spatially assigned to the 83 regions of the DK atlas94 using the abagen toolbox84,95, with regional assignment based on corrected Montreal National Institute Space coordinates. Assignments were constrained by hemisphere and cortical/subcortical boundaries to improve anatomical accuracy. Gene expression values were normalized using a robust sigmoid function and rescaled to a unit interval for cross-region and cross-donor comparability. Due to limited availability of samples from the right hemisphere, analyses were restricted to 41 left-hemisphere regions spanning 34 cortical and 7 subcortical divisions.
Large-scale networks
We assigned each of the DK atlas regions to 7 Yeo resting large-scale networks at rest96 and von Economo cytoarchitectonic classes97 following the procedure described in ref. 98. We then calculated mean PCA1 scores within each of these networks and classes to obtain a summary measure.
Developmental brain regional gene expression
The BrainSpan Atlas of the Developing Human Brain includes microarray and RNA-seq data from 42 neurotypical postmortem brains spanning key developmental stages, from 8 weeks post-conception to 40 years of age. Details of the methods underlying the generation of the dataset have been described elsewhere90. Briefly, microarray samples (GeneChip Human Exon 1.0 ST Array platform) measure the expression of transcript variants across over 50,000 genes and probe sets. Normalized gene expression data were downloaded for our candidate genes from https://www.brainspan.org/. For this analysis, microarray data were prioritized due to the extensive temporal and spatial coverage. Samples were organized into developmental stages: fetal (8–20 weeks post-conception), infancy (birth to 1 year), childhood (1–10 years), adolescence (11–20 years) and adulthood (21+ years). We averaged normalized gene expression across donors for the same region and developmental age before conducting further analyses.
PET templates
To integrate histaminergic gene expression with neurochemical profiles, we analyzed PET-derived maps of neurotransmitter receptor densities sourced from the neuromaps toolbox (https://netneurolab.github.io/neuromaps/). These templates included receptor binding measures for serotonin, dopamine, acetylcholine, GABA, glutamate and cannabinoid systems28. Datatsets from tracers for receptors of the histamine system were available for the H3 only: [11C]GSK189254 (ref. 99) and [11C]MK-8278 (ref. 100). These datasets provide in vivo regional maps of histamine H3 receptor availability in healthy adults and were selected because the H3 receptor acts as a key presynaptic autoreceptor regulating histamine synthesis and release, making it a particularly informative marker of histaminergic innervation at the systems level. Importantly, the inclusion of these PET templates allowed us to validate the spatial correspondence between postmortem histaminergic gene expression patterns and receptor distributions measured in the living human brain, thereby strengthening the biological plausibility of our multimodal findings. At present, histamine H1 receptor PET datasets are not available in a shareable form that allows reliable parcellation and integration with large-scale multimodal neuroimaging frameworks such as those used here. Neurotransmitter receptor densities were averaged within DK atlas regions to ensure alignment with the transcriptomic data (Supplementary Table 2).
Magnetoencephalography oscillatory neural dynamics maps
To examine whether our summary measure of histaminergic gene expression spatially align with the cortical distribution of oscillatory dynamics across different frequency ranges, we used parcellated data from magnetoencephalography maps acquired in healthy volunteers as part of the Human Connectome Project as in ref. 101. Briefly, the dataset pertains to 6 minutes of resting-state data acquired with the eyes open. Complete MEG acquisition protocols can be found in the HCP S1200 Release Manual102. For each participant, the power spectrum was calculated at the vertex level across six different frequency bands—delta (2–4 Hz), theta (5–7 Hz), alpha (8–12 Hz), beta (15–29 Hz), low gamma (30–59 Hz) and high gamma (60–90 Hz)—using the open-source software Brainstorm. This was implemented using the Welch’s method with overlapping windows of length 4 seconds with 50% overlap. Average power at each frequency band was then calculated for each vertex (that is, source). Source-level power data were then parcellated into the 68 cortical regions of the DK atlas for each frequency band to ensure alignment with the transcriptomics data.
Neurosynth meta-analytical data
Probabilistic measures of the association between voxels and terms were obtained from the Neurosynth repository, a meta-analytical tool that synthesizes results from more than 15,000 published functional MRI (fMRI) studies by searching for high-frequency key words (such as pain and attention) that are published alongside fMRI voxel coordinates (https://github.com/neurosynth/neurosynth, using the volumetric association test maps)88. This measure of association is the probability that a given term is reported in the study if there is activation observed at a given voxel. Note that the tool does not distinguish between areas that are activated or deactivated in relation to the term of interest, or the degree of activation, only that certain brain areas are frequently mentioned in conjunction with certain words. Although more than 1,000 terms are reported in Neurosynth, we followed the approach taken in ref. 98 and focused primarily on cognitive function, therefore limiting the terms of interest to cognitive and behavioral terms. These terms were selected from the Cognitive Atlas, a public ontology of cognitive science that includes a comprehensive list of neurocognitive terms and has previously been used in conjunction with Neurosynth. We used 123 terms, ranging from umbrella terms (attention and emotion) to specific cognitive processes (visual attention and episodic memory), behaviors (eating and sleep) and emotional states (fear and anxiety). The coordinates reported by Neurosynth were parcellated into regions of the DK atlas. The probabilistic measure reported by Neurosynth can be interpreted as a quantitative representation of how regional fluctuations in activity are related to psychological processes. For simplicity, we refer to these probabilities as functional associations throughout this paper. The full list of terms is shown in Supplementary Table 3.
ENIGMA structural brain changes in neuropsychiatric disorders
Meta-analytical data on the effect size of case–control structural differences in cortical thickness and surface area were obtained from the ENIGMA consortium for seven psychiatric disorders (ADHD, autism spectrum disorder, anorexia nervosa, bipolar disorder, MDD, obsessive–compulsive disorder and schizophrenia). These data are distributed as part of the ENIGMA toolbox89,103. A summary of the sample sizes and sociodemographic characteristics is provided in Supplementary Table 4.
Statistical analysis
All statistical analyses were implemented in Python and MATLAB using the assistance of OpenAI ChatGPT to help with debugging code errors when those occurred. Spatial correlations between maps were assessed with Spearman rank correlations using spin-based permutation testing (Vasa method104) to account for spatial autocorrelation. Gene expression was summarized using PCA implemented in Python, selecting the optimal number of components to retain using parallel simulations. For the developmental analyses, we used permutation tests shuffling samples across developmental age to assess the statistical significance of histaminergic gene expression trends over time. Corrections for multiple comparisons were conducted using the FDR to limit type I error rates. Plotting and data visualization were performed using Python’s Matplotlib (https://matplotlib.org/) and Seaborn (https://seaborn.pydata.org/) libraries and the ENIGMA toolbox89.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
All data used in this study are publicly available from established repositories. Postmortem human brain transcriptomic data were obtained from the Allen Human Brain Atlas (AHBA; https://human.brain-map.org). Developmental transcriptomic data were accessed from publicly available brain developmental atlases, including BrainSpan (https://www.brainspan.org). PET receptor templates were derived from previously published datasets and accessed via the neuromaps platform (https://neuromaps-main.readthedocs.io; https://github.com/netneurolab/neuromaps). Structural brain alteration maps for psychiatric disorders were obtained from the ENIGMA consortium (https://enigma.ini.usc.edu). Functional meta-analytic maps were accessed via Neurosynth (https://neurosynth.org). No new primary datasets were generated for this study. All processed data supporting the findings are available from the corresponding author on reasonable request, subject to the terms and conditions of the original data providers. This study exclusively used de-identified, publicly available datasets. All original data collection procedures were conducted by the respective data providers in accordance with institutional review board approvals and relevant ethical guidelines. No new human or animal participants were recruited, and no additional ethical approval was required for the present analyses.
Code availability
All analyses were conducted using custom scripts in Python and MATLAB, built on openly available toolboxes and resources. Mapping of Allen Human Brain Atlas microarray data to the Desikan–Killiany atlas was performed using abagen (https://abagen.readthedocs.io; https://github.com/rmarkello/abagen). Molecular PET templates were obtained and integrated via neuromaps (https://neuromaps-main.readthedocs.io; https://github.com/netneurolab/neuromaps). ENIGMA case–control structural effect-size maps were accessed and harmonized using the ENIGMA Toolbox (https://enigma-toolbox.readthedocs.io; https://github.com/MICA-MNI/ENIGMA). Meta-analytic functional decoding used Neurosynth association maps (https://neurosynth.org). MEG power spectra were processed in Brainstorm (https://neuroimage.usc.edu/brainstorm). Spatial autocorrelation-preserving permutation testing employed the spin-test implementation described by Vasa et al. (https://github.com/spin-test/spin-test). Principal component analysis and statistical analyses were implemented in Python using NumPy (https://numpy.org), SciPy (https://scipy.org) and scikit-learn (https://scikit-learn.org). Visualizations were produced with Matplotlib (https://matplotlib.org) and Seaborn (https://seaborn.pydata.org).
References
-
Bernardino, L. Histamine in the crosstalk between innate immune cells and neurons: relevance for brain homeostasis and disease. Curr. Top. Behav. Neurosci. 59, 261–288 (2022).
-
Carthy, E. & Ellender, T. Histamine, neuroinflammation and neurodevelopment: a review. Front. Neurosci. 15, 680214 (2021).
-
Nuutinen, S. & Panula, P. Histamine in neurotransmission and brain diseases. Adv. Exp. Med. Biol. 709, 95–107 (2010).
-
Wada, H. et al. Is the histaminergic neuron system a regulatory center for whole-brain activity?. Trends Neurosci. 14, 415–418 (1991).
-
Panula, P., Yang, H. Y. & Costa, E. Histamine-containing neurons in the rat hypothalamus. Proc. Natl. Acad. Sci. USA 81, 2572–2576 (1984).
-
Arrigoni, E. & Fuller, P. M. The role of the central histaminergic system in behavioral state control. Curr. Top. Behav. Neurosci. 59, 447–468 (2022).
-
Panula, P., Sundvik, M. & Karlstedt, K. Developmental roles of brain histamine. Trends Neurosci. 37, 159–168 (2014).
-
Xu, L. et al. The diverse network of brain histamine in feeding: dissect its functions in a circuit-specific way. Curr. Neuropharmacol. 22, 241–259 (2024).
-
Baronio, D. et al. Histaminergic system in brain disorders: lessons from the translational approach and future perspectives. Ann. Gen. Psychiatry 13, 34 (2014).
-
Onodera, K. et al. Neuropharmacology of the histaminergic neuron system in the brain and its relationship with behavioral disorders. Prog. Neurobiol. 42, 685–702 (1994).
-
Schwartz, J. C. et al. Histaminergic transmission in the mammalian brain. Physiol. Rev. 71, 1–51 (1991).
-
Gbahou, F., Rouleau, A. & Arrang, J. M. The histamine autoreceptor is a short isoform of the H(3) receptor. Br. J. Pharmacol. 166, 1860–1871 (2012).
-
Arrang, J. M., Garbarg, M. & Schwartz, J. C. Auto-inhibition of brain histamine release mediated by a novel class (H3) of histamine receptor. Nature 302, 832–837 (1983).
-
Peng, L. & Wang, T. Histamine synthesis and transport are coupled in axon terminals via a dual quality control system. EMBO J. 43, 4472–4491 (2024).
-
Yoshikawa, T., Nakamura, T. & Yanai, K. Histamine N-methyltransferase in the brain. Int. J. Mol. Sci. 20, 737 (2019).
-
Timmerman, H. Histamine receptors in the central nervous system. Pharm. Weekbl. Sci. 11, 146–150 (1989).
-
Watanabe, T. et al. Evidence for the presence of a histaminergic neuron system in the rat brain: an immunohistochemical analysis. Neurosci. Lett. 39, 249–254 (1983).
-
Futagawa, A. et al. Comprehensive mapping of histamine H(1) receptor mRNA in the mouse brain. J. Comp. Neurol. 532, e25622 (2024).
-
Inagaki, N. et al. Organization of histaminergic fibers in the rat brain. J. Comp. Neurol. 273, 283–300 (1988).
-
Panula, P. et al. Histamine-immunoreactive nerve fibers in the rat brain. Neuroscience 28, 585–610 (1989).
-
Airaksinen, M. S. & Panula, P. The histaminergic system in the guinea pig central nervous system: an immunocytochemical mapping study using an antiserum against histamine. J. Comp. Neurol. 273, 163–186 (1988).
-
Lin, W. et al. Whole-brain mapping of histaminergic projections in mouse brain. Proc. Natl. Acad. Sci. USA 120, e2216231120 (2023).
-
Flik, G. et al. Interaction between brain histamine and serotonin, norepinephrine, and dopamine systems: in vivo microdialysis and electrophysiology study. J. Mol. Neurosci. 56, 320–328 (2015).
-
Benoy, A. & Ramaswamy, S. Histamine in the neocortex: towards integrating multiscale effectors. Eur. J. Neurosci. 60, 4597–4623 (2024).
-
Haas, H. & Panula, P. The role of histamine and the tuberomamillary nucleus in the nervous system. Nat. Rev. Neurosci. 4, 121–130 (2003).
-
Yanai, K. & Tashiro, M. The physiological and pathophysiological roles of neuronal histamine: an insight from human positron emission tomography studies. Pharmacol. Ther. 113, 1–15 (2007).
-
Mochizuki, H. et al. Quantitative measurement of histamine H(1) receptors in human brains by PET and [11C]doxepin. Nucl. Med. Biol. 31, 165–171 (2004).
-
Markello, R. D. et al. neuromaps: structural and functional interpretation of brain maps. Nat. Methods 19, 1472–1479 (2022).
-
Ma, Q. et al. Histamine H(2) receptor deficit in glutamatergic neurons contributes to the pathogenesis of schizophrenia. Proc. Natl. Acad. Sci. USA 120, e2207003120 (2023).
-
Arias-Montano, J. A. et al. Histamine H(3) receptor-mediated inhibition of depolarization-induced, dopamine D(1) receptor-dependent release of [(3)H]-gamma-aminobutryic acid from rat striatal slices. Br. J. Pharmacol. 133, 165–171 (2001).
-
Serafim, K. R. et al. H1-histamine receptors in the amygdala are involved in emotional memory but do not mediate anxiety-related behaviors in mice submitted to EPM testing. Brain Res. Bull. 89, 1–7 (2012).
-
Gianlorenco, A. C. et al. Emotional memory consolidation impairment induced by histamine is mediated by H1 but not H2 receptors. Brain Res. Bull. 89, 197–202 (2012).
-
Schlicker, E. & Kathmann, M. Role of the histamine H(3) receptor in the central nervous system. Handb. Exp. Pharmacol. 241, 277–299 (2017).
-
Martins, D. et al. Imaging transcriptomics: convergent cellular, transcriptomic, and molecular neuroimaging signatures in the healthy adult human brain. Cell Rep. 37, 110173 (2021).
-
Sadek, B. et al. Histamine H3 receptor as a potential target for cognitive symptoms in neuropsychiatric diseases. Behav. Brain Res. 312, 415–430 (2016).
-
Vohora, D. & Bhowmik, M. Histamine H3 receptor antagonists/inverse agonists on cognitive and motor processes: relevance to Alzheimer’s disease, ADHD, schizophrenia, and drug abuse. Front. Syst. Neurosci. 6, 72 (2012).
-
Fukudo, S. et al. Histamine neuroimaging in stress-related disorders. Curr. Top. Behav. Neurosci. 59, 113–129 (2022).
-
Freitas, B. & Teodoro, T. P. Neuropsychiatry of histaminergic circuits: potential role of novel h3 receptor selective antagonist/inverse agonist pitolisant in Prader–Willi syndrome. Psychopharmacol. Bull. 54, 103–107 (2024).
-
Mercer, L. P. et al. Dietary induced anorexia: a review of involvement of the histaminergic system. J. Am. Coll. Nutr. 15, 223–230 (1996).
-
Schlicker, E. et al. Modulation of neurotransmitter release via histamine H3 heteroreceptors. Fundam. Clin. Pharmacol. 8, 128–137 (1994).
-
Schlicker, E., Betz, R. & Gothert, M. Histamine H3 receptor-mediated inhibition of serotonin release in the rat brain cortex. Naunyn Schmiedebergs Arch. Pharmacol. 337, 588–590 (1988).
-
Moreno, E. et al. Dopamine D1-histamine H3 receptor heteromers provide a selective link to MAPK signaling in GABAergic neurons of the direct striatal pathway. J. Biol. Chem. 286, 5846–5854 (2011).
-
Marquez-Gomez, R. et al. Functional histamine H(3) and adenosine A(2A) receptor heteromers in recombinant cells and rat striatum. Pharmacol. Res. 129, 515–525 (2018).
-
Ferrada, C. et al. Interactions between histamine H3 and dopamine D2 receptors and the implications for striatal function. Neuropharmacology 55, 190–197 (2008).
-
Ellender, T. J. et al. Differential modulation of excitatory and inhibitory striatal synaptic transmission by histamine. J. Neurosci. 31, 15340–15351 (2011).
-
Prast, H. et al. Histaminergic neurons modulate acetylcholine release in the ventral striatum: role of H3 histamine receptors. Naunyn Schmiedebergs Arch. Pharmacol. 360, 558–564 (1999).
-
Prast, H., Fischer, H. & Philippu, A. Release of acetylcholine in the ventral striatum is influenced by histamine receptors. Agents Actions 41, C85–C86 (1994).
-
Lin, J. S. et al. Involvement of histaminergic neurons in arousal mechanisms demonstrated with H3-receptor ligands in the cat. Brain Res. 523, 325–330 (1990).
-
Burgess, C. R. Histamine and orexin in the control of arousal, locomotion, and motivation. J. Neurosci. 30, 2810–2811 (2010).
-
Provensi, G. et al. Neuronal histamine and the memory of emotionally salient events. Br. J. Pharmacol. 177, 557–569 (2020).
-
Torrealba, F. et al. Histamine and motivation. Front. Syst. Neurosci. 6, 51 (2012).
-
Thakkar, M. M. Histamine in the regulation of wakefulness. Sleep Med. Rev. 15, 65–74 (2011).
-
Passani, M. B., Blandina, P. & Torrealba, F. The histamine H3 receptor and eating behavior. J. Pharmacol. Exp. Ther. 336, 24–29 (2011).
-
Mobarakeh, J. I. et al. Role of histamine H(1) receptor in pain perception: a study of the receptor gene knockout mice. Eur. J. Pharmacol. 391, 81–89 (2000).
-
Panula, P. & Nuutinen, S. The histaminergic network in the brain: basic organization and role in disease. Nat. Rev. Neurosci. 14, 472–487 (2013).
-
Kim, Y. J., Goto, Y. & Lee, Y. A. Histamine H3 receptor antagonists ameliorate attention deficit/hyperactivity disorder-like behavioral changes caused by neonatal habenula lesion. Behav. Pharmacol. 29, 71–78 (2018).
-
Provensi, G. et al. Brain histamine modulates recognition memory: possible implications in major cognitive disorders. Br. J. Pharmacol. 177, 539–556 (2020).
-
Kotanska, M. et al. Pitolisant protects mice chronically treated with corticosterone from some behavioral but not metabolic changes in corticosterone-induced depression model. Pharmacol. Biochem. Behav. 196, 172974 (2020).
-
Munari, L. et al. Brain histamine is crucial for selective serotonin reuptake inhibitors’ behavioral and neurochemical effects. Int. J. Neuropsychopharmacol. 18, pyv045 (2015).
-
Hersey, M. et al. Inflammation-induced histamine impairs the capacity of escitalopram to increase hippocampal extracellular serotonin. J. Neurosci. 41, 6564–6577 (2021).
-
Yang, C. et al. Inflammatory markers and treatment outcome in treatment resistant depression: a systematic review. J. Affect. Disord. 257, 640–649 (2019).
-
Arrang, J. M. Histamine and schizophrenia. Int. Rev. Neurobiol. 78, 247–287 (2007).
-
Arumuham, A. et al. Working memory processes and the histamine-3 receptor in schizophrenia: a [(11)C]MK-8278 PET-fMRI study. Psychopharmacology 242, 1321–1334 (2024).
-
Cheng, L. et al. Histamine H(1) receptor deletion in cholinergic neurons induces sensorimotor gating ability deficit and social impairments in mice. Nat. Commun. 12, 1142 (2021).
-
Meskanen, K. et al. A randomized clinical trial of histamine 2 receptor antagonism in treatment-resistant schizophrenia. J. Clin. Psychopharmacol. 33, 472–478 (2013).
-
Schwartz, J. C. The histamine H3 receptor: from discovery to clinical trials with pitolisant. Br. J. Pharmacol. 163, 713–721 (2011).
-
Passani, M. B. & Blandina, P. Histamine receptors in the CNS as targets for therapeutic intervention. Trends Pharmacol. Sci. 32, 242–249 (2011).
-
Yanai, K. et al. Histamine H1 receptors in human brain visualized in vivo by [11C]doxepin and positron emission tomography. Neurosci. Lett. 137, 145–148 (1992).
-
Qian, H. et al. Histamine and histamine receptors: roles in major depressive disorder. Front. Psychiatry 13, 825591 (2022).
-
Satpati, A., Neylan, T. & Grinberg, L. T. Histaminergic neurotransmission in aging and Alzheimer’s disease: a review of therapeutic opportunities and gaps. Alzheimers Dement. 9, e12379 (2023).
-
Yanai, K. et al. Age-dependent decrease in histamine H1 receptor in human brains revealed by PET. Neuroreport 3, 433–436 (1992).
-
Nakamura, T. et al. Brain histamine H(1) receptor occupancy after oral administration of desloratadine and loratadine. Pharmacol. Res. Perspect. 7, e00499 (2019).
-
Sarfraz, N. et al. Pitolisant, a novel histamine-3 receptor competitive antagonist, and inverse agonist, in the treatment of excessive daytime sleepiness in adult patients with narcolepsy. Health Psychol. Res. 10, 34222 (2022).
-
Shan, L. et al. Diurnal fluctuation in histidine decarboxylase expression, the rate limiting enzyme for histamine production, and its disorder in neurodegenerative diseases. Sleep 35, 713–715 (2012).
-
Yoshitake, T. et al. Determination of histamine in microdialysis samples from rat brain by microbore column liquid chromatography following intramolecular excimer-forming derivatization with pyrene-labeling reagent. J. Neurosci. Methods 127, 11–17 (2003).
-
Shen, E. H., Overly, C. C. & Jones, A. R. The Allen Human Brain Atlas: comprehensive gene expression mapping of the human brain. Trends Neurosci. 35, 711–714 (2012).
-
Piwecka, M., Rajewsky, N. & Rybak-Wolf, A. Single-cell and spatial transcriptomics: deciphering brain complexity in health and disease. Nat. Rev. Neurol. 19, 346–362 (2023).
-
Krejcar, O. & Namazi, H. Multiscale brain modeling: bridging microscopic and macroscopic brain dynamics for clinical and technological applications. Front. Cell. Neurosci. 19, 1537462 (2025).
-
Best, J. et al. A mathematical model for histamine synthesis, release, and control in varicosities. Theor. Biol. Med. Model. 14, 24 (2017).
-
Igaz, P. et al. Histamine genomics in silico: polymorphisms of the human genes involved in the synthesis, action and degradation of histamine. Am. J. Pharmacogenomics 2, 67–72 (2002).
-
Jones, B. L. et al. Genetic variation in the histamine production, response, and degradation pathway is associated with histamine pharmacodynamic response in children with asthma. Front. Pharmacol. 7, 524 (2016).
-
Micallef, S., Stark, H. & Sasse, A. Polymorphisms and genetic linkage of histamine receptors. Life Sci. 93, 487–494 (2013).
-
Hawrylycz, M. J. et al. An anatomically comprehensive atlas of the adult human brain transcriptome. Nature 489, 391–399 (2012).
-
Markello, R. D. et al. Standardizing workflows in imaging transcriptomics with the abagen toolbox. eLife 10, e72129 (2021).
-
Alexander, B. et al. Desikan–Killiany–Tourville atlas compatible version of M-CRIB neonatal parcellated whole brain atlas: the M-CRIB 2.0. Front. Neurosci. 13, 34 (2019).
-
Ashworth, S. et al. Evaluation of 11C-GSK189254 as a novel radioligand for the H3 receptor in humans using PET. J. Nucl. Med. 51, 1021–1029 (2010).
-
Kanba, S. & Richelson, E. Histamine H1 receptors in human brain labelled with [3H]doxepin. Brain Res. 304, 1–7 (1984).
-
Yarkoni, T. et al. Large-scale automated synthesis of human functional neuroimaging data. Nat. Methods 8, 665–670 (2011).
-
Lariviere, S. et al. The ENIGMA toolbox: multiscale neural contextualization of multisite neuroimaging datasets. Nat. Methods 18, 698–700 (2021).
-
Miller, J. A. et al. Transcriptional landscape of the prenatal human brain. Nature 508, 199–206 (2014).
-
Siletti, K. et al. Transcriptomic diversity of cell types across the adult human brain. Science 382, eadd7046 (2023).
-
Human Multiple Cortical Areas Smart-Seq (Allen Institute, 2026); https://portal.brain-map.org/atlases-and-data/rnaseq/human-multiple-cortical-areas-smart-seq
-
Arnatkeviciute, A., Fulcher, B. D. & Fornito, A. A practical guide to linking brain-wide gene expression and neuroimaging data. Neuroimage 189, 353–367 (2019).
-
Furtjes, A. E. et al. A quantified comparison of cortical atlases on the basis of trait morphometricity. Cortex 158, 110–126 (2023).
-
abagen: a toolbox for the Allen Brain Atlas genetics data (abagen, 2021); https://abagen.readthedocs.io/en/stable/
-
Yeo, B. T. et al. The organization of the human cerebral cortex estimated by intrinsic functional connectivity. J. Neurophysiol. 106, 1125–1165 (2011).
-
Triarhou, L. C. The Economo–Koskinas atlas revisited: cytoarchitectonics and functional context. Stereotact. Funct. Neurosurg. 85, 195–203 (2007).
-
Hansen, J. Y. et al. Mapping gene transcription and neurocognition across human neocortex. Nat. Hum. Behav. 5, 1240–1250 (2021).
-
Gallezot, J. D. et al. Determination of receptor occupancy in the presence of mass dose: [(11)C]GSK189254 PET imaging of histamine H(3) receptor occupancy by PF-03654746. J. Cereb. Blood Flow Metab. 37, 1095–1107 (2017).
-
Arumuham, A. et al. Working memory processes and the histamine-3 receptor in schizophrenia: a [(11)C]MK-8278 PET-fMRI study. Psychopharmacology 242, 1321–1334 (2025).
-
Hansen, J. Y. et al. Mapping neurotransmitter systems to the structural and functional organization of the human neocortex. Nat. Neurosci. 25, 1569–1581 (2022).
-
WU–Minn HCP 12 Subjects Data Release Reference Manual (WU–Minn Consortium Human Connectome Project, 2018); https://www.humanconnectome.org/storage/app/media/documentation/s1200/HCP_S1200_Release_Reference_Manual.pdf
-
ENIGMA Toolbox (enigmators, 2020); https://enigma-toolbox.readthedocs.io/en/latest/
-
Vasa, F. et al. Adolescent tuning of association cortex in human structural brain networks. Cereb. Cortex 28, 281–294 (2018).
-
Cell Types Database: RNA-Seq Data (Allen Institute, 2026); https://portal.brain-map.org/atlases-and-data/rnaseq
Acknowledgements
D.M. is supported by NIHR Maudsley Biomedical Research Centre, South London, and Maudsley NHS Trust.
Ethics declarations
Competing interests
The authors declare no competing interest.
Peer review
Peer review information
Nature Mental Health thanks Alex Doruyter, Maria Beatrice Passani and the other, anonymous, reviewer(s) for their contribution to the peer review of this work.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Martins, D., Veronese, M., van Wamelen, D. et al. Mapping histamine pathway networks in the human brain across cognition and psychiatric disorders. Nat. Mental Health 4, 816–828 (2026). https://doi.org/10.1038/s44220-026-00637-1
-
Received:
-
Accepted:
-
Published:
-
Version of record:
-
Issue date:
-
DOI: https://doi.org/10.1038/s44220-026-00637-1
Leave a Reply